
Dr William Stiles – Aberystwyth University
The technology of controlled environment agriculture (CEA) is fast developing and is increasingly recognised as an important component of future food production. A key advancement which has enhanced the potential for this approach to horticultural production has been the introduction of light emitting diode (LED) technology.
LEDs are much more efficient than previous horticultural lighting options, such as high-pressure sodium bulbs, both for the amount of energy consumed and the amount of photosynthetically active radiation (PAR) that can be delivered to the plant leaf. LEDs also offer huge flexibility with regard to the composition of light which is emitted, meaning that light can be tailored to specifically match the requirements of the plant, increasing efficiency. Additionally, and perhaps more intriguingly, light spectral composition can also be altered to induce novel responses in plants. In this way, growth form or the potential for phytochemical production, can be manipulated to achieve desired outcomes.
Light is an essential component of plant development and is a key driver of plant physiology and morphology. Light is crucial for photosynthesis, the chemical reaction that fixes CO2 for the purposes of food production, but it also acts as an environmental prompt, informing plants about the world in which they exist. Plants detect information from incoming light using sophisticated multi-functional sensory proteins called photoreceptors. Plants possess at least five classes of photoreceptor: phytochromes, cryptochromes, phototropins, Flavin-binding F-box proteins, and UVR8. Actual signalling pathways, and the interaction between different receptors, are complex, but in essence phytochromes perceive red and far-red light, cryptochromes perceive blue and UV-A light, phototropins and Flavin-binding F-box proteins blue light, and UVR8 perceives UV-B light.
Plants are organisms with high rates of plasticity allowing them to alter their physiology and development to adapt to changing environmental conditions, which is an essential strategy for sessile organisms that are unable to simply move to more favourable situations. The ability to vary light spectral composition, to favour or limit the proportion of certain wavebands, offers opportunities to optimise production approaches by mimicking environmental stimuli and triggering plant responses.
Light and photosynthesis Light is electromagnetic radiation that we perceive as visible light, within the wavelength range of 400-700 nanometres (nm). This ranges from ultraviolet light (with shorter wavelengths) to infrared light (with longer wavelengths). The electromagnetic spectrum also contains wavelengths of light which are invisible to the human eye, such as gamma rays, x-rays, microwaves and radio waves.
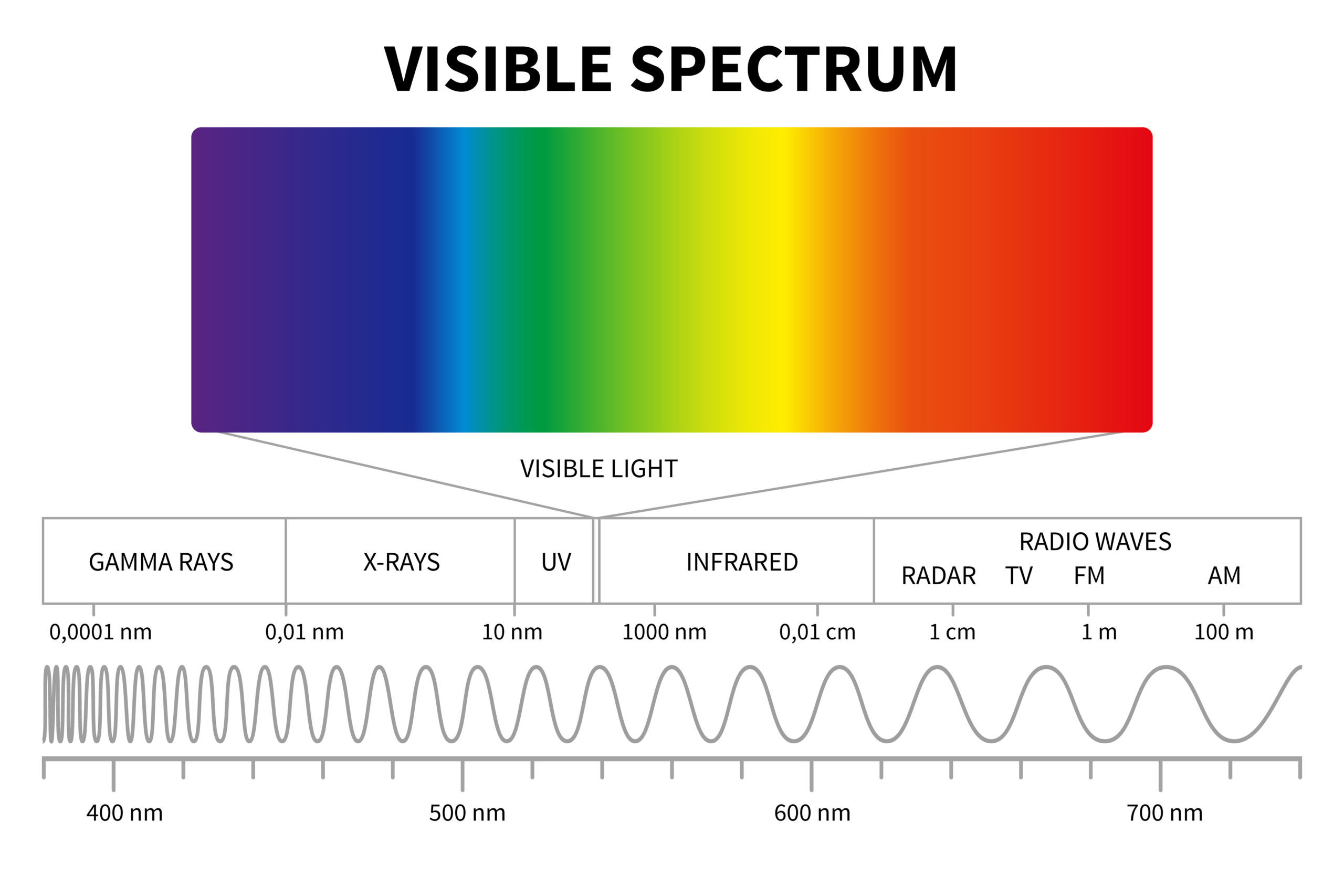
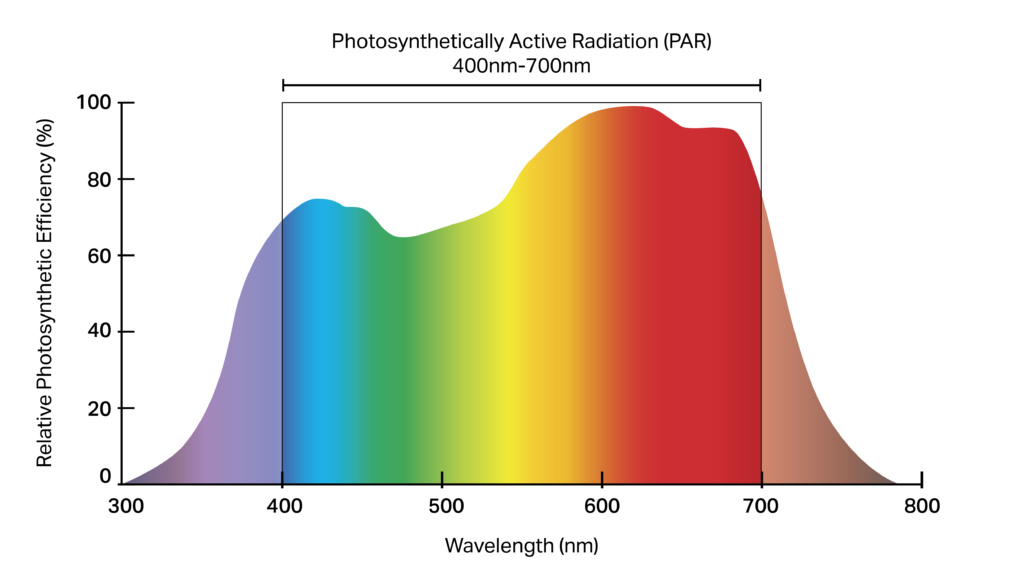
Light can be emitted from many different sources, including objects of sufficiently high temperature, but not all light sources are suitable for the purposes of photosynthesis. Light from within the visible spectrum drives photosynthesis, particularly light from within blue and red wavelength ranges, but the potential for photosynthesis will be governed by the amount of energy available in the form of photons that a plant can absorb. Light intensity, and its potential for driving photosynthesis, is referred to as the photosynthetic photon flux density (PPFD). The higher the PPFD, the higher the potential for photosynthesis. Plants absorb light energy via the light-absorbing pigment chlorophyll. Chlorophyll appears green as it absorbs all visible light except green wavelengths, which are reflected. Chlorophyll A and B absorb red
and blue light strongly, and as such these wavebands have been considered the only portion of light that truly matters for plant production. However, increasingly it is recognised that plants make use of all available light to at least some degree, including green light, and that presence (or absence) of different wavebands influences plant development.
Light effects on plants
Every plant has a high degree of plasticity with regard to response to environmental conditions. This means it will look very different, and grow very differently, depending on the characteristics of an environment. Where light availability and composition influence plant development and growth patterns, this is referred to as photomorphogenesis. There are three stages of development where photomorphogenesis affects development: seed germination, seedling development, and the switch from vegetative to flowering stage.
In this way light composition can exert strong influence over plant growth form. Understanding the influence of light in development is key for promoting growth that is favourable for a controlled environment. For instance, plants grown in red spectrum light will exhibit greater leaf curling, and plants grown with exposure to blue spectrum light will exhibit greater leaf flattening. This may mean that a high rate of blue could make plants better able to utilise light which is unidirectional and only (at least primarily) coming from above, typical for controlled growing scenarios. Of course there are numerous other considerations, and potential ramifications, to consider, but this does at least highlight that factors such as this are key considerations in a controlled environment where the selection of light spectra may mean that the individual responses from the plant may actually make the plant more, or less, suited to growing in that environment.
Each of the wavebands of the light spectrum, and their relative proportion in the available light, will trigger a response in the plant. The different wavebands are:
Red light (600-700 nm) – light from the red wavelengths is the main driver of vegetative growth. This means more leaves and more biomass. But growing in the absence of other spectra may result in a phenomena referred to as red-light syndrome, where leaf photosynthesis can become impaired. Without the presence of blue light, the form or morphology of plant tissues may also result in unfavourable growth profiles, where plants become stretched and tall, with thin leaves, which is a typically unfavourable growth profile. It may also mean plants cannot utilise all available light energy, leading to overall inefficiency. Overall, red is the most important wavelength for plant growth and development, but not in isolation.
Blue light (400-500 nm) – light from the blue portion of the spectrum has a large effect on plant morphology. It can increase the ratio of root to shoot in plant development, promoting root growth and plant compactness, which has certain implications depending on production goals. Blue light also promotes more stomatal opening, which means more stomatal conductance and gas exchange. This is typically considered favourable from a plant health perspective but may result in greater humidity potential, which is a consideration for controlled environments. Blue light is absorbed readily by plant photoreceptors, and is an important factor in plant environmental perception. For instance, increasing the percentage of blue light will convince plants that there is more available light overall, which will change plant behaviour.
Green light (500-600 nm) – green light is weakly absorbed compared to red and blue wavelengths, but is increasingly recognised as important for overall photosynthesis potential. Green light is reflected and scattered within leaves and the canopy, which increases the potential for total absorption. Green is particularly important in dense-growing scenarios where there is a large amount of shading, as it drives photosynthesis in lower or shaded leaves. Green light also affects morphology via the green to blue light ratio. This acts as a signal to indicate shade conditions, informing the plant and leaf of its position in the canopy, initiating growth behaviour associated with shade avoidance. This can include extra growth or stretching of the internode and leaf length, and the angle of the leaves may also change to capture more incidental rather than direct light.
Far-red (700-850 nm) – this portion of light is referred to as super-visual, as the majority of this waveband is outside the visible portion of the spectrum. Far-red is not considered conventionally photosynthetically active and it only weakly drives photosynthesis, but adding far-red will change how plants grow as this light is absorbed by phytochrome photoreceptors, which are involved in the regulation of leaf expansion, flowering, internode extension, and the partitioning of resources between organs. Far-red will also have the opposite effect to blue light on root to shoot ratio, resulting in higher shoot to root distribution. Yet, as with all elements of the light spectrum, there is a balance to be struck between a beneficial amount of far-red light and too much. Plants grown under high levels of far-red light will appear tall and stretched, with lower chlorophyll content resulting in yellowing of the leaves, which is perhaps unfavourable from a marketability perspective. In addition to direct effects, the ratio of red to far-red light is also an important mechanism for governing plant responses. Far-red penetrates the canopy more than red light, so plants receiving a higher amount of far-red relative to red will interpret this as a shading effect, and increase shade avoidance responses such as increased upwards growth.
UV spectrum (100-400 nm) – UV light is also outside of the PAR wavelengths, but this light will still affect plant development. Plant responses to UV-A light are similar to blue light. UV-B is higher energy and has its own photo receptor in plants, called UVR8. Adding UV-B to the spectrum will change the morphology in ways which are not considered essential for survival, but which may affect the potential for production. For instance, under UV-B light plant cuticles can grow thicker, making the plants generally more robust, and UV-B exposure will positively regulate stomatal development, but hypocotyl and petiole length may be shorter and rosette leaf expansion may be impaired. Secondary metabolite production is also higher under UV-B, which is typically a favourable response, particularly for production systems focussing on pharmaceutical production. UV-C is not believed to be directly perceived by plants, but it can be highly useful for the control of pests and disease in controlled environments.
Light influence on other organisms
In addition to the influence different light spectrum compositions will have on plants in controlled production systems, different light spectrums may also influence other organisms, including those considered pests or pathogens.
Light intensity, photoperiod, and wavelength composition will affect certain pest and pathogen organism behaviours. This can offer the potential for both positive and negative effects, as the right light composition may make it challenging for pests and pathogens to persist, or alternatively may offer optimum conditions for them to thrive. Different light wavelengths can influence invertebrate/insect perceptions and reproduction rate; for instance, UV light is key for insect sight, which is effective in the UV-A region, making the identification of target plants easier for insect herbivores, and UV-B and UV-C light can have a detrimental effect on insect and fungi populations, reducing their impact. Additionally, some interaction between plants and pest organisms may also be affected by the provision of certain light wavelengths, such as far-red, where light induced responses in plants such as phenolic compound production may improve plant natural defences against herbivorous invertebrates.
The degree to which this is an issue will depend to some degree on the production system. In hyper-controlled scenarios, it is possible that invertebrate herbivores may be totally excluded, yet nevertheless some consideration of unintended effects of light spectrum choice is necessary to ensure problematic production challenges are avoided where possible. Nevertheless, finding a perfect balance between light requirements for the plant and light for pest management is likely to be highly challenging as these two objectives are unlikely to be aligned.
Summary
As LED lighting technology develops it is increasingly possible to alter the composition of light, to select individual wavebands or to alter the proportion of specific wavelengths, in order to elicit a targeted growth response in plants. The ability to manipulate the composition of light, and an understanding of the biological mechanisms at the heart of plant responses to different light compositions and availabilities, can therefore be used to ‘instruct’ plants as to how you want them to grow.
The motivations for doing this are varied, but the principal reasons will either be to increase the amount of biomass produced, and therefore overall yield, to alter the shape and appearance of the plant, or change its chemical composition, to make it more appealing to a customer and therefore more commercially attractive.
The selection of spectra can be complex however, as including, or excluding, light from a given wavelength will have both direct effects and indirect effects. Nevertheless, manipulating light in this way offers the horticultural sector many options to optimise growth regimes, or to determine recipes for light provision that can result in novel structures, or bio-compound production, that may offer enormous commercial advantage.